BIOLOGÍA CELULAR
CITOLOGÍA
Kytos: Célula
Logos: Estudio
CITOLOGÍA: Es
una disciplina académica que se encarga del estudio de las células en cuanto a
lo que respecta a las propiedades, estructura, funciones, orgánulos que
contienen, su interacción con el ambiente y su ciclo vital.
CÉLULA:
Unidad funcional y estructural de todo ser vivo.
Las células poseen membrana, citoplasma, material genético (ADN) y (ARN), éste último puede ser: ARN mensajero (ARNm), ARN de transferencia (ARNt), ARN ribosómico (ARNr).
Unidad funcional y estructural de todo ser vivo.
Las células poseen membrana, citoplasma, material genético (ADN) y (ARN), éste último puede ser: ARN mensajero (ARNm), ARN de transferencia (ARNt), ARN ribosómico (ARNr).

POSTULADOS

AÑO
|
PERSONAJE
|
ACONTECIMIENTO
|
1665
|
Robert Hooke
|
Construyó un microscopio y observó que los tejidos
vegetales estaban formados por pequeñas celdas a las que llamo células.
|
1676
|
Antonio Van
Leeuwenhoek
|
Construyó
microscopios de mayor aumento, descubriendo así la existencia de los
microorganismos.
|
1831
|
Robert Brown
|
Observó que el núcleo estaba en todas las celular
vegetales.
|
1838
|
Theodor Shwann
|
Postuló
que la célula era un principio de construcción de organismos más complejos.
|
1855
|
Remarck y Virchow
|
Afirmaron que toda célula proviene de otra célula.
|
1865
|
Gregor Mendel
|
Establece
dos principios genéticos:
|
1869
|
Friedrich Miescher
|
Aisló el Ácido Desoxirribonucleico (AND).
|
1902
|
Suttony Boveri
|
Refiere
que la información biológica hereditaria reside en los cromosomas.
|
1911
|
Sturtevant
|
Comenzó a construir mapas cromosómicos donde observó los locus y los locis de los
genes.
|
1914
|
Robert Feulgen
|
Descubrió
que el ADN podía teñirse con fucsina, demostrando que el ADN se encuentra en
los cromosomas de todas las células.
|
1953
|
Watson y Crick
|
Elaboraron un modelo de la doble hélice del ADN.
|
1997
|
Ian Wilmut
|
Científico
que clono a la oveja Dolly.
|
2000
|
Estados Unidos, Gran Bretaña,
Francia, Alemania, Japón y China
|
Las investigaciones realizadas por estos países
dieron lugar al primer borrador del Genoma Humano. Actualmente el mapa del
genoma humano esta descubierto en un 85 %.
|
MITOSIS Y MEISOSIS
EL MICROSCOPIO
Es
un instrumento que permite observar objetos que son demasiado pequeños para ser
vistos a simple vista. El tipo más común y el primero que se inventó es el
microscopio óptico. Se trata de un instrumento óptico que contiene dos o más
lentes que permiten obtener una imagen aumentada del objeto y que funciona por
refracción. La ciencia que investiga los objetos pequeños utilizando este
instrumento se llama microscopía.
Inventado por:
Zacharias
Janssen en 1590. En 1665 aparece en la obra de William Harvey sobre la circulación
sanguínea al mirar al
microscopio los capilares
sanguíneos y Robert Hooke publica su obra Micrographia.
En
1665 Robert
Hooke observó con un
microscopio un delgado corte de corcho
y notó que el material era poroso, en su conjunto, formaban cavidades poco
profundas a modo de celditas a las que llamó células. Se trataba de la primera observación
de células muertas. Unos años más tarde, Marcello Malpighi, anatomista y biólogo italiano,
observó células vivas. Fue el primero en estudiar tejidos vivos al microscopio.
A
mediados del siglo XVII un holandés, Anton
van Leeuwenhoek,
utilizando microscopios simples de fabricación propia, describió por primera
vez protozoos, bacterias, espermatozoides y glóbulos rojos. El microscopista Leeuwenhoek, sin
ninguna preparación científica, puede considerarse el fundador de la bacteriología. Tallaba él mismo sus lupas, sobre
pequeñas esferas de cristal, cuyos diámetros no alcanzaban el milímetro (su
campo de visión era muy limitado, de décimas de milímetro). Con estas pequeñas
distancias focales alcanzaba los 275 aumentos. Observó los glóbulos de la
sangre, las bacterias y los protozoos; examinó por primera vez los glóbulos
rojos y descubrió que el semen contiene espermatozoides. Durante su
vida no reveló sus métodos secretos y a su muerte, en 1723,
26 de sus aparatos fueron cedidos a la Royal
Society de Londres.
Durante
el siglo XVIII continuó el progreso y se lograron objetivos acromáticos por
asociación de Chris
Neros y Flint
Crown obtenidos en
1740 por H. M. Hall y mejorados por John Dollond. De esta época son los estudios
efectuados por Isaac
Newton y Leonhard Euler. En el siglo XIX, al descubrirse que
la dispersión y la refracción se podían modificar con combinaciones
adecuadas de dos o más medios ópticos, se lanzan al mercado objetivos
acromáticos excelentes.
CÉLULA EUCARIOTA ANIMAL

ESTRUCTURA
DE LAS EUCARIOTAS
MEMBRANA PLASMATICA
Es el límite externo de la célula, que le da protección y actúa como una
barrera selectiva entre el líquido del espacio extracelular y el citoplasma. La
composición de la membrana plasmática incluye alrededor de un 40 % de lípidos y
50 % proteínas, junto a pequeñas cantidades de hidratos de carbono, cerca del
10 %, unidas a las dos anteriores. Los lípidos están representados por una
doble capa de fosfolípidos y por otros lípidos como el colesterol, este último
solo en eucariotas animales. La formación de la bicapa se debe a que los
fosfolípidos son anfipáticos, es decir, cada molécula posee una región
hidrofílica, soluble en agua, y una región hidrofóbica que repele el agua. Las
cabezas hidrofílicas se orientan hacia el citoplasma y hacia el medio
extracelular, mientras que las colas hidrófobas lo hacen hacia el interior de
la membrana. Del total de lípidos que conforman la membrana plasmática, cerca
del 75 % son fosfolípidos, mientras que el 20 % corresponden a moléculas de
colesterol. Estos últimos brindan mayor fortaleza a la membrana y la hacen
menos deformable. Además, el colesterol disminuye la permeabilidad de la bicapa
a pequeñas moléculas hidrosolubles. El 5 % restante de los lípidos de membrana
son los glucolípidos, elementos unidos a los lípidos con características
anfipáticas que se orientan hacia el área extracelular. Los glucolípidos
contribuyen a mantener la adhesión de células y tejidos.
Membrana plasmática
{kind=link}
Dentro del 50 % de las proteínas que conforman la membrana plasmática hay
diferentes tipos. Las denominadas proteínas integrales se unen fuertemente a
los lípidos y atraviesan la doble capa. La mayor parte de las proteínas
integrales son glucoproteínas, donde el monosacárido se orienta al medio
extracelular. Las proteínas periféricas se asocian débilmente a los lípidos y
se ubican a uno u otro lado de la membrana, sin atravesarla, en contacto con
las cabezas hidrófilas de los fosfolípidos.
Esquema de
un sector de la membrana plasmática
{kind=link}
Las proteínas de la membrana plasmática tienen funciones de comunicación,
de unión a receptores moleculares, de transporte, de acción enzimática, de
anclaje de filamentos del citoesqueleto y de identidad celular, entre otros. Es
así que las proteínas de la membrana plasmática:
- Establecen canales a manera de poros por donde entran y salen sustancias
de la célula, siendo las proteínas integrales las encargadas de esa misión.
- Reconocen y se unen a receptores de ciertas moléculas que sean
importantes para la célula, como nutrientes, hormonas, neurotransmisores, etc.
Esta función también la realizan las proteínas integrales.
- Transportan sustancias a través de la membrana plasmática.
- Poseen acción enzimática en la superficie de la membrana, catalizando reacciones bioquímicas. De esta función se encargan las proteínas integrales y periféricas
- Transportan sustancias a través de la membrana plasmática.
- Poseen acción enzimática en la superficie de la membrana, catalizando reacciones bioquímicas. De esta función se encargan las proteínas integrales y periféricas
- Fijan los filamentos del citoesqueleto celular, a cargo de ambos tipos de
proteínas.
- Regulan la identidad celular por medio de las glucoproteínas y
glucolípidos, haciendo posible identificar a las células que provienen de otros
organismos, como sucede en las transfusiones de sangre. Los glóbulos rojos, al
presentar marcadores del tipo A, B, AB y O permiten identificar sangres
compatibles. Es decir, la determinación del tipo de sangre en humanos se
relaciona con la clase de glucolípidos existente en la superficie de los
eritrocitos. Las porciones hidrocarbonadas de las glucoproteínas y los
glucolípidos forman el glucocálix, que actúa como una capa protectora.
Las membranas de los distintos organoides del citoplasma son las encargadas de mantener las diferentes concentraciones de sustancias que hay en el interior de las estructuras y en el citosol. El grosor aproximado de la membrana plasmática es de 70 - 80 Å (angstrom). Un ángstrom es igual a la diez millonésima parte de un milímetro (1 Å = 0,0000001 mm).
Las membranas de los distintos organoides del citoplasma son las encargadas de mantener las diferentes concentraciones de sustancias que hay en el interior de las estructuras y en el citosol. El grosor aproximado de la membrana plasmática es de 70 - 80 Å (angstrom). Un ángstrom es igual a la diez millonésima parte de un milímetro (1 Å = 0,0000001 mm).
La membrana plasmática se une a las membranas plasmáticas de las células
vecinas mediante estructuras engrosadas llamadas desmosomas. De función
mecánica, los desmosomas mantienen a las células bien ligadas entre sí, por
medio de filamentos proteicos anclados al citoesqueleto. Estos parches son
abundantes en el tejido epitelial y en el músculo cardíaco, áreas sujetas a una
importante tensión mecánica.
{kind=link}
CITOPLASMA
Es la parte de la célula que se ubica entre la membrana plasmática y la membrana nuclear. Está constituido por 85 % de agua y un 15 % de proteínas, aminoácidos, sales y minerales. En el citoplasma se realizan la mayoría de las reacciones metabólicas de la célula.
La porción del citoplasma sin estructura y que forma la parte fluida se denomina hialoplasma o citosol, lugar donde están las moléculas necesarias para el mantenimiento de la célula. Vale decir que el hialoplasma es el medio interno líquido de todas las estructuras celulares.
El citoesqueleto es una serie de filamentos proteicos responsable de la forma celular y de facilitar el movimiento de los organoides. Actúa como una conexión entre las distintas partes de la célula. El citoesqueleto se destruye y se vuelve a reconstruir, por lo que no es una estructura permanente de la célula. Se forma a partir de tres componentes proteicos: microtúbulos, microfilamentos y filamentos intermedios.
- Microtúbulos: son los componentes más importantes del citoesqueleto, compuestos por una proteína denominada “tubulina”. De consistencia rígida, son los responsables de la formación de estructuras como los centríolos y órganos de locomoción, como los cilios y los flagelos. Los microtúbulos irradian desde el centrosoma.
- Microfilamentos: se disponen cerca de la membrana plasmática y están asociados al movimiento de la célula. Están formados por dos tipos de proteínas, la “actina” y la “miosina”. Los microfilamentos están muy desarrollados en células musculares estriadas (músculos voluntarios). La superposición de microfilamentos de actina y miosina permiten la contracción muscular.
Es la parte de la célula que se ubica entre la membrana plasmática y la membrana nuclear. Está constituido por 85 % de agua y un 15 % de proteínas, aminoácidos, sales y minerales. En el citoplasma se realizan la mayoría de las reacciones metabólicas de la célula.
La porción del citoplasma sin estructura y que forma la parte fluida se denomina hialoplasma o citosol, lugar donde están las moléculas necesarias para el mantenimiento de la célula. Vale decir que el hialoplasma es el medio interno líquido de todas las estructuras celulares.
El citoesqueleto es una serie de filamentos proteicos responsable de la forma celular y de facilitar el movimiento de los organoides. Actúa como una conexión entre las distintas partes de la célula. El citoesqueleto se destruye y se vuelve a reconstruir, por lo que no es una estructura permanente de la célula. Se forma a partir de tres componentes proteicos: microtúbulos, microfilamentos y filamentos intermedios.
- Microtúbulos: son los componentes más importantes del citoesqueleto, compuestos por una proteína denominada “tubulina”. De consistencia rígida, son los responsables de la formación de estructuras como los centríolos y órganos de locomoción, como los cilios y los flagelos. Los microtúbulos irradian desde el centrosoma.
- Microfilamentos: se disponen cerca de la membrana plasmática y están asociados al movimiento de la célula. Están formados por dos tipos de proteínas, la “actina” y la “miosina”. Los microfilamentos están muy desarrollados en células musculares estriadas (músculos voluntarios). La superposición de microfilamentos de actina y miosina permiten la contracción muscular.
- Filamentos intermedios: están formados por varios tipos de
proteínas. Se extienden por todo el citoplasma y abundan en aquellas células
que soportan mucha tensión, por lo que son resistentes y evitan la destrucción
celular.
{kind=link}
Dentro del citoplasma, existen organelas con distintas funciones, que están
presentes tanto en eucariotas animales como vegetales y que se detallan a
continuación.
MITOCONDRIAS
Las mitocondrias son organelas que presentan doble membrana, una externa en contacto con el citoplasma y otra interna, hacia la matriz mitocondrial. Dicha matriz está compuesta por agua y proteínas. Las mitocondrias, de forma oval y alargada, son consideradas como las “usinas eléctricas” de las células. Son las encargadas de producir y almacenar energía en forma de ATP a partir de la glucosa, lípidos y demás nutrientes. Mediante la respiración celular, proceso que consume oxígeno y libera dióxido de carbono, se produce energía que se acumula en el ATP. Toda vez que en algún lugar de la célula se necesita aporte energético, por ejemplo para transportar sustancias a través de la membrana plasmática, la división celular, reciclado de desechos, etc., el ATP se descompone y se libera.
Las mitocondrias poseen ADN en su interior, un ARN propio y ribosomas. Las mitocondrias ocupan un lugar importante dentro del citoplasma. Algunas células del organismo con una actividad energética importante, como las hepáticas y las musculares, poseen gran cantidad de mitocondrias por cada célula.
{kind=link}
RETICULO ENDOPLASMÁTICO
Esta estructura es un sistema de membranas que se dispone formando una red
de sacos aplanados, donde contiene túbulos que se conectan entre sí formando
una lámina continua que da lugar a un lumen. Las membranas del retículo
endoplásmico separan dicho lumen del citoplasma, y actúan en la transferencia
selectiva de moléculas entre ambos compartimientos. Todas las membranas del retículo endoplasmático equivalen a la mitad de las
membranas totales que hay en cada célula. Además, es el lugar donde se producen
todas las proteínas y los lípidos que forman las membranas del propio retículo,
del complejo de Golgi, de los lisosomas y de la membrana plasmática.
El retículo endoplásmico adopta dos variedades: una forma granular o rugosa
y otra agranular o lisa. El retículo endoplasmático granular está unido a la
membrana nuclear externa, mientras que el retículo endoplasmático agranular es
una prolongación del retículo endoplasmático rugoso.
Son funciones del retículo la síntesis de proteínas, de lípidos, el
transporte intracelular de sustancias y la detoxificación de la célula. Las
sustancias sintetizadas son almacenadas y luego transportadas a su destino
celular. La detoxificación de sustancias como fármacos, drogas y desechos
celulares es de gran importancia en las células del hígado.
{kind=link}
a)
Retículo endoplasmático granular (REG)
Presenta numerosas protuberancias, debido a una gran cantidad de ribosomas
unidos a la membrana. Tiene por función la detoxificación celular y la síntesis
de proteínas que serán utilizadas por las membranas, por otras organelas o para
ser enviadas fuera de la célula. Estas membranas forman un espacio interno (luz
del retículo) que lo separan del citoplasma, condicionando la transferencia
selectiva de moléculas entre ambos compartimientos.
El REG tiene gran desarrollo en células del hígado y del páncreas, debido a una intensa labor detoxificante y de síntesis. También en los glóbulos blancos, ya que producen y secretan anticuerpos (proteínas) para ser exportadas a todo el organismo.
El REG tiene gran desarrollo en células del hígado y del páncreas, debido a una intensa labor detoxificante y de síntesis. También en los glóbulos blancos, ya que producen y secretan anticuerpos (proteínas) para ser exportadas a todo el organismo.
b)
Retículo endoplasmático agranular (REA)
Su apariencia “lisa” se debe a la carencia de ribosomas. Dentro del REA
están las enzimas necesarias para la síntesis de lípidos (triglicéridos,
fosfolípidos y esteroides) y enzimas necesarias para la detoxificación de
alcoholes y otras sustancias. El REA es importante de células del testículo y
del ovario para la síntesis de hormonas esteroides, como también en los
hepatocitos para detoxificar sustancias nocivas.
{kind=link}
COMPLEJO DE GOLGI
Es un
organoide con 5 a 10 sacos aplanados membranosos de forma discoide denominados
dictiosomas. Estos dictiosomas se conectan entre sí y contienen fluidos en su
interior. Poseen una cara cóncava y otra convexa. La parte cóncava (cara cis o
de formación), próxima al retículo endoplásmico, recibe de este último las
proteínas sintetizadas en el área rugosa (granular). Esas proteínas son
transportadas en vesículas de transición hasta la mencionada cara cis del
complejo de Golgi. La parte convexa del dictiosoma (cara trans o de maduración)
es la más cercana a la membrana plasmática y
formadora de vesículas de secreción. En síntesis, las vesículas de transición
que llegan del retículo endoplásmico penetran en la cara cis del complejo de
Golgi, atraviesan todos los sáculos o dictiosomas y llegan al trans-Golgi. Aquí
son empaquetadas para luego dirigirse a la membrana plasmática para vaciar su
contenido fuera de la célula por exocitosis.
El complejo de Golgi secreta sustancias tales como enzimas digestivas, hormonas y sustancias que se transforman en glucoproteínas para la formación de la pared celular. Algunas enzimas permanecen dentro de vesículas membranosas, los lisosomas, capaces de degradar moléculas complejas. El complejo de Golgi se encuentra en todas las células eucariotas, a excepción de las epidérmicas y de los glóbulos rojos.
El complejo de Golgi secreta sustancias tales como enzimas digestivas, hormonas y sustancias que se transforman en glucoproteínas para la formación de la pared celular. Algunas enzimas permanecen dentro de vesículas membranosas, los lisosomas, capaces de degradar moléculas complejas. El complejo de Golgi se encuentra en todas las células eucariotas, a excepción de las epidérmicas y de los glóbulos rojos.
En resumen, las funciones del complejo de Golgi son:
-Síntesis de polisacáridos para la pared celular.
-Formación de glucoproteínas y glucolípidos de secreción. (Glicosilación de prótidos y lípidos).
-Síntesis de polisacáridos para la pared celular.
-Formación de glucoproteínas y glucolípidos de secreción. (Glicosilación de prótidos y lípidos).
-Formación
de lisosomas que permanecen en el citoplasma.
-Empaquetamiento, dentro de vesículas, de sustancias de secreción como proteínas. - Transporte intracelular de sustancias.
-Empaquetamiento, dentro de vesículas, de sustancias de secreción como proteínas. - Transporte intracelular de sustancias.
{kind=link}
RIBOSOMAS
Son organelas muy pequeñas de alrededor de 20 nanómetros de diámetro, visibles al microscopio electrónico. Un nanómetro es la millonésima parte del milímetro (1 nm = 0,000001 mm).
Los ribosomas están formados por ácido ribonucleico ribosómico (ARNr) y proteínas. Existen varios millones de estas estructuras en cada célula. Los ribosomas se componen de dos subunidades: una mayor, que se encarga de formar las uniones de aminoácidos que darán lugar a las proteínas, y otra menor que reconoce a los ARN mensajeros (ARNm) y a los ARN de transferencia (ARNt). En eucariotas, las dos subunidades mencionadas se sintetizan en el nucléolo. Las moléculas de ARN mensajero llevan la información que llega desde el ADN de cómo se distribuirán los aminoácidos para la elaboración de una determinada proteína. El ARN de transferencia transporta los aminoácidos apropiados hacia los ribosomas para que se incorporen a las proteínas.
Son organelas muy pequeñas de alrededor de 20 nanómetros de diámetro, visibles al microscopio electrónico. Un nanómetro es la millonésima parte del milímetro (1 nm = 0,000001 mm).
Los ribosomas están formados por ácido ribonucleico ribosómico (ARNr) y proteínas. Existen varios millones de estas estructuras en cada célula. Los ribosomas se componen de dos subunidades: una mayor, que se encarga de formar las uniones de aminoácidos que darán lugar a las proteínas, y otra menor que reconoce a los ARN mensajeros (ARNm) y a los ARN de transferencia (ARNt). En eucariotas, las dos subunidades mencionadas se sintetizan en el nucléolo. Las moléculas de ARN mensajero llevan la información que llega desde el ADN de cómo se distribuirán los aminoácidos para la elaboración de una determinada proteína. El ARN de transferencia transporta los aminoácidos apropiados hacia los ribosomas para que se incorporen a las proteínas.
Hay ribosomas que permanecen libres en el citoplasma, fuera del retículo endoplásmico, cuya misión es sintetizar proteínas que permanecen dentro de las células. Los ribosomas de células procariotas y eucariotas tienen la misma forma y función. Se sintetizan dentro del núcleo celular, más precisamente en el nucléolo.
{kind=link}
NUCLEO
Esta la
organelo, que ocupa la parte central de la célula, actúa como centro de control
de casi toda la actividad hereditaria. Está delimitado por dos membranas
concéntricas, la carioteca o envoltura nuclear, donde se distinguen una
membrana externa que contacta con el citoplasma y otra interna. Sobre esta
última, hay una delgada capa de filamentos proteicos, la lámina nuclear, que
actúa fijando a los cromosomas cuando la célula se divide.
La carioteca, que se conecta de manera directa con el retículo endoplásmico, posee numerosos orificios o poros por donde salen hacia el citoplasma moléculas de ARN mensajero que serán leídas por los ribosomas, proteínas y ARN ribosómico, precursor de los ribosomas. Las sustancias que ingresan por los poros del citoplasma al núcleo son proteínas sintetizadas por los ribosomas citoplasmáticos. Los poros nucleares regulan en forma selectiva el pasaje de sustancias. Una de las funciones de la carioteca es proteger al ADN intranuclear de las distintas reacciones que se producen en el citoplasma.
El jugo nuclear es una sustancia que llena todo el núcleo, formada por una solución coloidal que contiene agua, carbohidratos, enzimas y ATP, entre otros.
Dentro del núcleo y en íntimo contacto con el jugo nuclear se encuentra la cromatina, que son filamentos muy largos y numerosos de ADN que se enrollan a moléculas de proteínas especiales llamadas “histonas”. Toda vez que una célula inicia su división, los filamentos de ADN se pliegan entre sí dando lugar a la formación de cromosomas. En un determinado lugar de los cromosomas se ubican los genes, que son subdivisiones o porciones de ADN. Es decir, cada gen es una secuencia de ADN que almacena información que se transmite a la descendencia.
Otra formación presente dentro del núcleo es el nucléolo, pequeña estructura de forma redondeada y sin membranas. Cuando las células comienzan a reproducirse (mitosis) el nucléolo desaparece, haciéndose nuevamente visible al final de la mitosis. El nucléolo contiene ADN ribosómico, fundamental para el proceso de fabricación de ARN (transcripción), que ha de sintetizar los ribosomas del citoplasma. Se ha comprobado que el nucléolo actúa como un regulador del ciclo celular.
La carioteca, que se conecta de manera directa con el retículo endoplásmico, posee numerosos orificios o poros por donde salen hacia el citoplasma moléculas de ARN mensajero que serán leídas por los ribosomas, proteínas y ARN ribosómico, precursor de los ribosomas. Las sustancias que ingresan por los poros del citoplasma al núcleo son proteínas sintetizadas por los ribosomas citoplasmáticos. Los poros nucleares regulan en forma selectiva el pasaje de sustancias. Una de las funciones de la carioteca es proteger al ADN intranuclear de las distintas reacciones que se producen en el citoplasma.
El jugo nuclear es una sustancia que llena todo el núcleo, formada por una solución coloidal que contiene agua, carbohidratos, enzimas y ATP, entre otros.
Dentro del núcleo y en íntimo contacto con el jugo nuclear se encuentra la cromatina, que son filamentos muy largos y numerosos de ADN que se enrollan a moléculas de proteínas especiales llamadas “histonas”. Toda vez que una célula inicia su división, los filamentos de ADN se pliegan entre sí dando lugar a la formación de cromosomas. En un determinado lugar de los cromosomas se ubican los genes, que son subdivisiones o porciones de ADN. Es decir, cada gen es una secuencia de ADN que almacena información que se transmite a la descendencia.
Otra formación presente dentro del núcleo es el nucléolo, pequeña estructura de forma redondeada y sin membranas. Cuando las células comienzan a reproducirse (mitosis) el nucléolo desaparece, haciéndose nuevamente visible al final de la mitosis. El nucléolo contiene ADN ribosómico, fundamental para el proceso de fabricación de ARN (transcripción), que ha de sintetizar los ribosomas del citoplasma. Se ha comprobado que el nucléolo actúa como un regulador del ciclo celular.
{kind=link}
Dentro de las importantes funciones que tiene el núcleo sobresale la de
ejercer el control de todas las actividades de la célula. Además, interviene en
la replicación y transcripción de los ácidos nucleicos y en la transferencia de
la información genética a las células hijas durante la división celular.
{kind=link}
I-
ESTRUCTURAS DE EUCARIOTAS ANIMALES
Las células eucariotas animales tienen estructuras exclusivas, como el centrosoma, los lisosomas y los cilios y flagelos. Además, se diferencian de las eucariotas vegetales por ser heterótrofas, por carecer de pared celular y de plástidos, estos últimos fundamentales para que los vegetales verdes puedan fotosintetizar.
LISOSOMAS
Los lisosomas se originan en los dictiosomas (sacos aplanados) del aparato de Golgi, y en ocasiones a partir de vesículas en algunas regiones del retículo endoplasmático granular. Son organelas pequeñas, esféricas y semejantes a vacuolas.
Limitadas por una sola membrana, contienen en su interior poderosas enzimas encargadas de digerir sustancias que ingresan a las células (lisosomas digestivos), con lo cual se comportan como un sistema digestivo celular. Por otra parte, los lisosomas pueden degradar desechos celulares, lípidos y proteínas (lisosomas autofágicos) que son liberados a través de la membrana plasmática.
Las células eucariotas animales tienen estructuras exclusivas, como el centrosoma, los lisosomas y los cilios y flagelos. Además, se diferencian de las eucariotas vegetales por ser heterótrofas, por carecer de pared celular y de plástidos, estos últimos fundamentales para que los vegetales verdes puedan fotosintetizar.
LISOSOMAS
Los lisosomas se originan en los dictiosomas (sacos aplanados) del aparato de Golgi, y en ocasiones a partir de vesículas en algunas regiones del retículo endoplasmático granular. Son organelas pequeñas, esféricas y semejantes a vacuolas.
Limitadas por una sola membrana, contienen en su interior poderosas enzimas encargadas de digerir sustancias que ingresan a las células (lisosomas digestivos), con lo cual se comportan como un sistema digestivo celular. Por otra parte, los lisosomas pueden degradar desechos celulares, lípidos y proteínas (lisosomas autofágicos) que son liberados a través de la membrana plasmática.
{kind=link}
Mecanismo de acción de los lisosomas
{kind=link}
CENTROSOMA
Ocupa un área del citoplasma situada casi siempre muy cerca del núcleo. Regula los movimientos celulares de cilios y flagelos y tiene un rol fundamental en la división celular. El centrosoma está formado por el diplosoma, la centrosfera y el áster. En su interior está el diplosoma, que son dos cilindros huecos cuyas paredes están formadas por unidades de proteína (figura de la izquierda). Esos cilindros son los centríolos, que carecen de membranas y se ubican de manera perpendicular entre sí. Cada célula posee dos centríolos, cuya función es intervenir en la división celular y posibilitar la transferencia de material genético entre las células hijas. Por cada centríolo hay nueve grupos de tres microtúbulos cada uno, dispuestos en forma cilíndrica.
Los centríolos se hacen visibles toda vez que la célula se divide para
reproducirse. La centrosfera es una sustancia traslúcida donde se ubica el
diplosoma. El áster es el conjunto de filamentos radiales que parten de la
centrosfera, fundamentales en el proceso de la mitosis.
{kind=link}
CILIOS Y FLAGELOS
Son proyecciones del citoesqueleto limitadas por una membrana que es
continuación de la membrana plasmática. Son estructuras similares y
permanentes. Los flagelos se caracterizan por ser largos y escasos. Los cilios por ser
cortos y numerosos. Dentro del citoplasma, ambos
están formados por un anillo representado por nueve pares de microtúbulos que
rodean a un par ubicado en el centro, todo cubierto por la membrana plasmática.
Muchas eucariotas, igual que las procariotas bacterianas, utilizan estas
estructuras para la locomoción. Son ejemplos el flagelo (cola) de los
espermatozoides y los cilios del paramecio.
{kind=link}
Los cilios (del latín “pestaña”) son prolongaciones muy finas de
la membrana plasmática a modo de “dedo de guante”, con un contenido que es
continuación del citoplasma. De diámetro uniforme en toda su longitud, rodean
total o parcialmente el contorno de las células. Los cilios producen
vibraciones sincronizadas que permiten el movimiento de la célula. El Paramecio
es un ejemplo de microorganismo ciliado, con cerca de 200 cilios en cada
individuo. Los cilios de las células del tracto respiratorio tienen la misión
de capturar las partículas del aire.
{kind=link}
Los flagelos son apéndices en forma de látigo presente en muchos
organismos unicelulares, como el Trypanosoma y en algunos pluricelulares, como
los espermatozoides. El flagelo es utilizado para la movilidad celular en
medios líquidos, igual que los cilios. También poseen un diámetro uniforme en
toda su longitud, aunque algo mayor. Son más largos y menos numerosos que los
cilios, ya que algunas células tienen tan solo uno o dos flagelos. El flagelo
de las eucariotas se desplaza como si fuera un látigo, mientras que en las
procariotas el movimiento es rotatorio a manera de sacacorcho.
CÉLULA EUCARIOTA VEGETAL

II-
ESTRUCTURAS DE EUCARIOTAS VEGETALES
A pesar que las eucariotas vegetales tienen casi los mismos elementos que
las eucariotas animales, hay estructuras que son propias como la pared celular
y los plástidos.
PARED CELULAR
Es una típica estructura de eucariotas vegetales y fúngicas. Se ubica en la
parte externa de la membrana plasmática, en contacto con células adyacentes. Es
de consistencia gruesa y rígida, formada principalmente por celulosa. Cumple
una función similar al esqueleto de los animales superiores, ya que le da
firmeza a la planta posibilitando que se mantenga erguida. Además, interviene
en diversos procesos como la absorción, secreción, transpiración y defensa
contra agentes patógenos. La pared está perforada por pequeños poros
denominados plasmodesmas. Estos plasmodesmas atraviesan la membrana plasmática
y establecen una comunicación directa entre el citoplasma de las células
adyacentes.
En la pared celular se diferencian tres estructuras, desde el exterior
hacia la parte interna de la célula. Ellas son: una lámina media, una pared
primaria y una pared secundaria, esta última en contacto con la membrana
plasmática y subdividida en tres capas. La celulosa de las paredes celulares
protege a las células adyacentes de la desecación. Gran parte de la corteza y
de la madera de los árboles está formada de pared celular.
{kind=link}
PLASTIDOS
Tal como las mitocondrias, los plástidos son organelas con doble membrana, responsables de los diferentes colores que tienen las plantas. Dentro de los plástidos, también llamados “plastos”, se distinguen los cloroplastos, los cromoplastos y los leucoplastos. Los cloroplastos contienen clorofila, que se encarga de captar la energía lumínica y transformarla en energía química. De esa forma, el vegetal realiza la fotosíntesis, reacción que tiene lugar en los tilacoides, sacos o vesículas aplanadas que están inmersos en una solución llamada estroma en el interior de los cloroplastos. En la membrana de los tilacoides se ubica la clorofila, carotenos y xantinas. Pilas de tilacoides forman el grana de los cloroplastos. Los cloroplastos producen grande cantidades de ATP (adenosintrifosfato). Contienen ADN, un ARN propio y ribosomas.
Los cromoplastos fabrican y almacenan otros pigmentos que le dan color a los frutos, flores y hojas secas. Son ejemplos de esos pigmentos el caroteno (anaranjado) y la xantofila (amarillo). Los leucoplastos son plástidos de color blanquecino encargados de almacenar almidones (amiloplastos), lípidos y proteínas.
Tal como las mitocondrias, los plástidos son organelas con doble membrana, responsables de los diferentes colores que tienen las plantas. Dentro de los plástidos, también llamados “plastos”, se distinguen los cloroplastos, los cromoplastos y los leucoplastos. Los cloroplastos contienen clorofila, que se encarga de captar la energía lumínica y transformarla en energía química. De esa forma, el vegetal realiza la fotosíntesis, reacción que tiene lugar en los tilacoides, sacos o vesículas aplanadas que están inmersos en una solución llamada estroma en el interior de los cloroplastos. En la membrana de los tilacoides se ubica la clorofila, carotenos y xantinas. Pilas de tilacoides forman el grana de los cloroplastos. Los cloroplastos producen grande cantidades de ATP (adenosintrifosfato). Contienen ADN, un ARN propio y ribosomas.
Los cromoplastos fabrican y almacenan otros pigmentos que le dan color a los frutos, flores y hojas secas. Son ejemplos de esos pigmentos el caroteno (anaranjado) y la xantofila (amarillo). Los leucoplastos son plástidos de color blanquecino encargados de almacenar almidones (amiloplastos), lípidos y proteínas.
{kind=link}
VACUOLAS
Son elementos en forma de saco que se originan a partir de provacuolas, pequeñas estructuras presentes en células jóvenes. A medida que la célula crece, estas diminutas estructuras absorben agua por ósmosis y se unen entre sí hasta formar una vacuola de gran tamaño que ocupa un considerable espacio del citoplasma. Las vacuolas tienen una membrana de permeabilidad selectiva que acumula agua, dando lugar al crecimiento de la célula y al mantenimiento de su turgencia. En su interior contiene sales, glúcidos, proteínas y demás nutrientes.
Son elementos en forma de saco que se originan a partir de provacuolas, pequeñas estructuras presentes en células jóvenes. A medida que la célula crece, estas diminutas estructuras absorben agua por ósmosis y se unen entre sí hasta formar una vacuola de gran tamaño que ocupa un considerable espacio del citoplasma. Las vacuolas tienen una membrana de permeabilidad selectiva que acumula agua, dando lugar al crecimiento de la célula y al mantenimiento de su turgencia. En su interior contiene sales, glúcidos, proteínas y demás nutrientes.
Las vacuolas también actúan en la remoción de elementos innecesarios.
Mediante el proceso de exocitosis (movimiento de sustancias hacia fuera de la
célula) las vacuolas se acercan y se adhieren a la membrana plasmática para
eliminar desechos al exterior. Además, por endocitosis (movimiento de
sustancias hacia dentro de la célula) pueden transportar al citoplasma
moléculas que no difunden por la membrana celular. En este caso, esas moléculas
se adhieren a la membrana plasmática y se produce una invaginación, formándose
una vacuola.
CÉLULA PROCARIOTA BACTERIANA

LAS BACTERIAS
Son células sin núcleo, la zona de la célula, donde
está el ADN y ARN no está limitado por membrana. Ej. Bacteria.
Actualmente están divididas en dos grupos:
• Eubacterias, que poseen paredes celulares
formadas por peptidoglicano o por mureína. Incluye a la mayoría de las
bacterias y también a las cianobacterias.
• Arqueobacterias, que utilizan otras sustancias para constituir sus paredes celulares. Son todas aquellas características que habitan en condiciones extremas como manantiales sulfurosos calientes o aguas de salinidad muy elevada.
• Arqueobacterias, que utilizan otras sustancias para constituir sus paredes celulares. Son todas aquellas características que habitan en condiciones extremas como manantiales sulfurosos calientes o aguas de salinidad muy elevada.
BIOLOGÍA
ESTUDIANTE: Katherine
Marisol Aguilar Marquez.
PROFESOR: Bioq.
Carlos García MsC.
TEMA: Síntesis
de proteínas celular.
OBJETIVO:
Explicar en qué consiste la transcripción y traducción.
SÍNTESIS DE PROTEÍNAS CELULARES.
Se
conoce como síntesis de proteínas
al proceso por el cual se componen nuevas proteínas a partir de los veinte aminoácidos
esenciales. En este
proceso, se transcribe el ADN en ARN. La síntesis de proteínas se realiza en
los ribosomas situados en el citoplasma celular.
En
el proceso de síntesis, los aminoácidos son transportados por
ARN de transferencia correspondiente para cada aminoácido hasta el ARN
mensajero donde se unen en la posición adecuada para formar las nuevas
proteínas.
TRANSCRIPCIÓN

La transcripción
del ADN es el primer proceso de la expresión génica, mediante el cual se
transfiere la información contenida en la secuencia del ADN hacia la secuencia
de proteína utilizando diversos ARN como intermediarios. Durante la
transcripción genética, las secuencias de ADN son copiadas a ARN mediante una
enzima llamada ARN polimerasa que sintetiza un ARN mensajero que mantiene la
información de la secuencia del ADN. De esta manera, la transcripción del ADN
también podría llamarse síntesis del ARN mensajero.
Transcripción
en eucariotas
En el caso de las eucariotas, el proceso se realiza en el núcleo, y es
similar al de las procariotas, pero de mayor complejidad. Diferentes ARNp
transcriben distintos tipos de genes. La ARNpII transcribe los pre-ARNm,
mientras que la ARNpI y ARNpIII transcriben los ARN-ribosomales y ARNt,
respectivamente. Los ARNs transcritos son modificados posteriormente. El pre-ARNm,
por ejemplo, sufre un proceso de maduración que tras cortes y empalmes
sucesivos elimina ciertos segmentos del ADN llamados los intrones para
producir el ARNm final. Durante este proceso de maduración se puede dar lugar a
diferentes moléculas de ARN, en función de diversos reguladores. Así pues, un
mismo gen o secuencia de ADN, puede dar lugar a diferentes moléculas de ARNm y
por tanto, producir diferentes proteínas. Otro factor de regulación propio de
las células eucariotas son los conocidos «potenciadores» que incrementan mucho
(100 veces) la actividad de transcripción de un gen, y no depende de la
ubicación de éstos en el gen, ni la dirección de la lectura.
Etapas de la
transcripción
Clásicamente se divide el proceso de la transcripción
en 3 etapas principales (iniciación, elongación y terminación), pero realmente
se pueden diferenciar 5 etapas:
Preiniciación
Al contrario de la replicación de ADN, durante el inicio de la transcripción no se requiere
la presencia de un cebador para sintetizar la nueva cadena, de ARN en este
caso. Antes del inicio de la transcripción se necesita toda una serie de factores de transcripción que ejercen los factores de iniciación. Estos se unen
a secuencias específicas de ADN para reconocer el sitio donde la transcripción
ha de comenzar. Esta secuencia de ADN en la que se ensamblan los complejos de
transcripción se llama promotor. Los
promotores se localizan en los extremos 5'-terminales de los genes, antes del comienzo del gen, y a ellos se unen los factores de transcripción mediante fuerzas de Van der Waals y enlaces de hidrógeno. Los promotores tienen secuencias reguladoras definidas, muy conservadas en cada especie, donde las
más conocidas son la caja TATA (situada
sobre la región -10), con la secuencia consenso TATA(A/T)A(A/T); y la caja
TTGACA (situada en el punto -35).
La formación del complejo de transcripción se
realiza sobre el promotor TATA, allí se forma el núcleo del complejo de
iniciación. Sobre la caja TATA se fija una proteína de unión (TBP) junto con el
factor de transcripción TFII D (TF proviene del inglés: transcription factor). Después, a
ellos se unen otros factores de transcripción específicos: TFII B se
une a TBP, TFII A (opcional), que estabiliza el complejo TFII B-TBP;
luego se une el complejo TFII F y ARN polimerasa, y al final TFII
E y TFII H. Todo ello forma un complejo que se llama «complejo
de preiniciación cerrado» o PIC. Cuando la estructura se abre por mediación del
factor de transcripción TFII H, da comienzo la iniciación y al
«complejo abierto» (por su acción helicasa dependiente
de ATP).
Iniciación
Primero, una Helicasa separa las hebras de ADN en
estas denominadas cajas TATA, ya que entre adenina y timina se establecen dos
enlaces de hidrógeno, mientras que entre citosina y guanina se forman tres.
Posteriormente se unen los factores y las proteínas de transcripción (TBP,
TF2D, TF2B) permitiendo, de esta manera, el acceso de la ARN polimerasa al
molde de ADN de cadena simple, siendo esta la última en posicionarse. Aunque la
búsqueda del promotor por la ARN polimerasa es muy rápida, la formación de la
«burbuja de transcripción» o apertura del ADN y la síntesis del cebador es muy
lenta. La burbuja de transcripción es una apertura de ADN desnaturalizado de 18
pares de bases, donde empieza a sintetizarse el ARN cebador a partir del
nucleótido número 10 del ADN molde de la burbuja de transcripción. La burbuja
de transcripción se llama «complejo abierto».
La ARN polimerasa es una enzima
formada por 5 subunidades: 2 subunidades α, 1 subunidad β, 1 subunidad β' y 1
subunidad ω que tiene como función la unión de ribonucleótidos trifosfato. Cuando se
forma el complejo abierto, la ARN polimerasa comienza a unir ribonucleótidos
mediante enlaces fosfodiéster, y una vez
que se forma el primer enlace fosfodiéster, acaba la etapa de iniciación y
comienza así la siguiente etapa.
Disgregación
del promotor
Una vez sintetizado el primer enlace fosfodiéster, se
debe deshacer el complejo del promotor para que quede limpio para volver a
funcionar de nuevo. Durante esta fase hay una tendencia a desprenderse el
transcrito inicial de ARN y producir transcritos truncados, dando lugar a una
iniciación abortada, común tanto en procariontes como eucariontes. Una vez que
la cadena transcrita alcanza una longitud de unos 23 nucleótidos, el complejo
ya no se desliza y da lugar a la siguiente fase, la elongación.
La disgregación del promotor coincide con una
fosforilación de la serina 5 del dominio carboxilo terminal de la ARN
polimerasa, que es fosforilado por el TFII H (que es una proteína quinasa dependiente
de ATP).
Elongación
La ARN polimerasa cataliza la elongación de cadena del
ARN. Una cadena de ARN se une por apareamiento de bases a la cadena de ADN, y
para que se formen correctamente los enlaces de hidrógeno que determina el
siguiente nucleótido del molde de ADN, el centro activo de la ARN polimerasa
reconoce a los ribo nucleótidos trifosfato entrantes. Cuando el nucleótido
entrante forma los enlaces de hidrógeno idóneos, entonces la ARN polimerasa
cataliza la formación del enlace fosfodiéster que corresponde. A esto se le llama elongación, la
segunda etapa de la transcripción del ARN.
Terminación
Al finalizar la síntesis de ARNm, esta molécula ya se
ha separado completamente del ADN (que recupera su forma original) y también de
la ARN polimerasa, terminando la transcripción. La terminación es otra etapa
distinta de la transcripción, porque justo cuando el complejo de transcripción
se ha ensamblado activamente debe desensamblarse una vez que la elongación se
ha completado. La terminación está señalizada por la información contenida en
sitios de la secuencia del ADN que se está transcribiendo, por lo que la ARN
polimerasa se detiene al transcribir algunas secuencias especiales del ADN.
Estas secuencias son ricas en guanina y citosina, situadas en el extremo de los
genes, seguidas de secuencias ricas en timina, formando secuencias palindrómicas, que cuando
se transcriben el ARN recién sintetizado adopta una «estructura en horquilla»
que desestabiliza el complejo ARN-ADN, obligando a separarse de la ARN
polimerasa, renaturalizándose la burbuja de transcripción. Algunas secuencias
de ADN carecen de la secuencia de terminación, sino que poseen otra secuencia a
la que se unen una serie de proteínas reguladoras específicas de la terminación
de la transcripción como rho.
TRADUCCIÓN

El ARN mensajero es el que lleva la información para
la síntesis de proteínas, es decir, determina el orden en que se unirán los
aminoácidos.
Esta información está codificada en forma de
tripletes, cada tres bases constituyen un codón que determina un aminoácido.
Las reglas de correspondencia entre codones y aminoácidos constituyen el código genético.
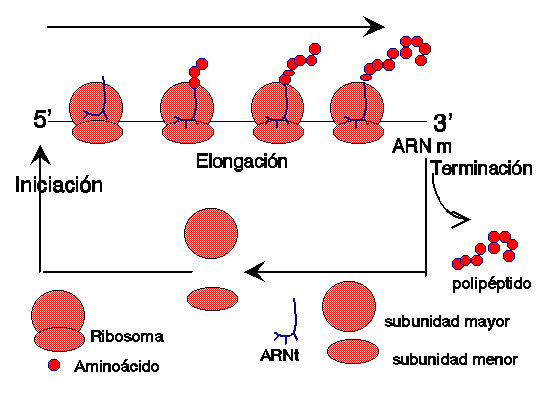
La síntesis de proteínas o traducción tiene lugar en los ribosomas del citoplasma. Los
aminoácidos son transportados por el ARN de transferencia, específico para cada
uno de ellos, y son llevados hasta el ARN mensajero, dónde se aparean el codón
de éste y el anticodón del ARN de transferencia, por complementariedad de
bases, y de ésta forma se sitúan en la posición que les corresponde.
Una vez finalizada la síntesis de una proteína, el ARN
mensajero queda libre y puede ser leído de nuevo. De hecho, es muy frecuente
que antes de que finalice una proteína
ya está comenzando otra, con lo cual, una misma molécula de ARN
mensajero, está siendo utilizada por varios
ribosomas simultanéamente.
 |
En esta maqueta se ha representado
el ARN mensajero como una varilla con los
codones (juego de tres colores). El ribosoma está
fijado al filamento, y las moléculas de ARN
transferencia, con los anticodones unidos a los codones del ARNm . En
la parte superior se observan tres aminoácidos unidos.
|
DIVISIÓN CELULAR
MITOSIS
La división celular es una parte muy importante del ciclo celular en la que una célula inicial se divide para formar células hijas. Gracias a la división celular se produce el crecimiento de los seres
vivos. En los organismos pluricelulares este crecimiento se produce
gracias al desarrollo de los tejidos y en los seres unicelulares mediante la reproducción vegetativa
.
Los seres pluricelulares reemplazan su dotación celular gracias a la división celular y suele estar asociada con la diferenciación celular.
En algunos animales la división celular se detiene en algún momento y
las células acaban envejeciendo. Las células senescentes se deterioran y
mueren debido al envejecimiento del cuerpo. Las células dejan de
dividirse porque los telómeros se vuelven cada vez más cortos en cada
división y no pueden proteger a los cromosomas como tal.
MITOSIS
La
mitosis es un proceso que ocurre
en el núcleo de las células eucarióticas y que precede inmediatamente a la división celular, consistente en el reparto
equitativo del material hereditario (ADN) característico. Este
tipo de división ocurre en las células somáticas y normalmente concluye con la
formación de dos núcleos separados (cariocinesis),
seguido de la partición del citoplasma (citocinesis),
para formar dos células hijas.
La
mitosis completa, que produce células genéticamente idénticas, es el fundamento
del crecimiento, de la reparación tisular y de la reproducción asexual. La otra forma de
división del material genético de un núcleo se denomina meiosis
y es un proceso que, aunque comparte mecanismos con la mitosis, no debe
confundirse con ella ya que es propio de la división celular de los gametos.
Produce células genéticamente distintas y, combinada con la fecundación, es el
fundamento de la reproducción sexual y la variabilidad genética.
Importancia de la Mitosis:
- Garantiza que las células hijas conserven el mismo número de cromosomas que la célula madre.
- Algunos seres unicelulares se reproducen asexualmente y de esta forma perpetúan la especie.
- Otros organismos reparan tejidos.
- Permite que se transmitan los caracteres hereditarios, ya que estas células poseen las unidades básicas de la herencia, los genes, que tienen toda la información necesaria para regular el desarrollo y las actividades metabólicas de la célula.

Profase: pro: primero, antes.
El centriolo
se duplica y emigra a los polos de la célula, alrededor de éste aparecen una
fibrillas llamadas áster. En las células vegetales se forma una estructura
semejante llamada Casquete Polar. Esta área se llama centrosoma.
Se forma el
huso acromático, una serie de filamentos que unen los centrosomas.
Desaparece el
huso y la membrana nuclear.
Al separarse
las cromátidas se mantienen unidas por el centrómero.

Metafase: meta: después, entre.
Termina por
desintegrarse la membrana nuclear.
Los
cromosomas se colocan en el plano ecuatorial de la célula.
El centrómero
de cada cromosoma entra en contacto con el huso acromático, se divide y las dos
cromátidas se separan en cromosomas idénticos.

Anafase: ana: arriba, ascendente.
Las
cromátidas se separan en sus centrómeros, y un juego de cromosomas se dirigen
hacia los polos de la célula.

Telofase: telos: fin
Se inicia
cuando los cromosomas ya están ubicados en los polos, entonces se alargan y
empiezan a desenrollarse.
El huso
acromático desaparece y se forman las membranas nucleares, cada una conserva el
número original de cromosomas. Se organizan los nuevos núcleos y reaparecen los
nucléolos.
Durante la
mitosis ocurren dos etapas muy importantes para la formación de las células
hijas:

Cariocinesis:
Es la división del núcleo celular.
Citocinesis:
Es la división del citoplasma
La Citocinesis:
Ocurre de
forma diferente en las células animales y vegetales. En la célula animal, desde
la anafase, cuando los cromosomas emigran a los polos se empieza a formar un
surco en el citoplasma, el cual se concentra en el ecuador celular. En la
telofase, el surco se hace más profundo hasta que se separa en dos mitades con
idéntico
contenido genético que la célula progenitora. En la célula vegetal, durante la
anafase, se forma una placa celular o tabique. Una vez organizado el material
genético, la placa origina a cada lado de ella una pared celular, la cual
termina por reparar las células hijas.
MEIOSIS
Meiosis es una de las formas de la reproducción celular. Este proceso se realiza en las glándulas sexuales para la producción de gametos. Es un proceso de división celular en el cual una célula diploide (2n) experimenta dos divisiones sucesivas, con la capacidad de generar cuatro células haploides (n). En los organismos con reproducción sexual tiene importancia ya que es el mecanismo por el que se producen los óvulos y espermatozoides (gametos).
Este proceso se lleva a cabo en dos divisiones nucleares y
citoplasmáticas, llamadas primera y segunda división meiótica o
simplemente meiosis I y meiosis II. Ambas comprenden profase, metafase, anafase y telofase.

Durante la meiosis los miembros de cada par homólogo de cromosomas se emparejan durante la profase, formando bivalentes. Durante esta fase se forma una estructura proteica denominada complejo sinaptonémico,
permitiendo que se produzca la recombinación entre ambos cromosomas
homólogos. Posteriormente se produce una gran condensación cromosómica y
los bivalentes se sitúan en la placa ecuatorial durante la primera
metafase, dando lugar a la migración de n cromosomas a cada uno
de los polos durante la primera anafase. Esta división reduccional es la
responsable del mantenimiento del número cromosómico característico de
cada especie. En la meiosis II, las cromátidas hermanas que forman cada
cromosoma se separan y se distribuyen entre los núcleos de las células
hijas. Entre estas dos etapas sucesivas no existe la etapa S
(replicación del ADN). La maduración de las células hijas dará lugar a los gametos.
Profase I

La Profase I de la primera división meiótica es la etapa más compleja del proceso y a su vez se divide en 5 subetapas, que son:
- Leptoteno
La primera etapa de Profase I es la etapa del leptoteno,
durante la cual los cromosomas individuales comienzan a condensar en
filamentos largos dentro del núcleo. Cada cromosoma tiene un elemento axial,
un armazón proteico que lo recorre a lo largo, y por el cual se ancla a
la envuelta nuclear. A lo largo de los cromosomas van apareciendo unos
pequeños engrosamientos denominados cromómeros. La masa cromática es 4c y es diploide 2n.
- Zigoteno
Los cromosomas homólogos comienzan a acercarse hasta quedar recombinados en toda su longitud. Esto se conoce como sinapsis (unión) y el complejo resultante se conoce como bivalente o tétrada (nombre que prefieren los citogenetistas),
donde los cromosomas homólogos (paterno y materno) se aparean,
asociándose así cromátidas homólogas. Producto de la sinapsis, se forma
el complejo sinaptonémico (estructura observable solo con el microscopio electrónico).
La disposición de los cromómeros
a lo largo del cromosoma parece estar determinado genéticamente. Tal es
así que incluso se utiliza la disposición de estos cromómeros para
poder distinguir cada cromosoma durante la profase I meiótica.
Además el eje proteico central pasa a formar los elementos laterales
del complejo sinaptonémico, una estructura proteica con forma de
escalera formada por dos elementos laterales y uno central que se van
cerrando a modo de cremallera y que garantiza el perfecto apareamiento
entre homólogos. En el apareamiento entre homólogos también está
implicada la secuencia de genes de cada cromosoma, lo cual evita el
apareamiento entre cromosomas no homólogos.
Durante el zigoteno concluye la replicación del ADN (2% restante) que recibe el nombre de zig-ADN.
- Paquiteno
Una vez que los cromosomas homólogos están perfectamente apareados
formando estructuras que se denominan bivalentes se produce el fenómeno
de entrecruzamiento cromosómico (crossing-over)
en el cual las cromátidas homólogas no hermanas intercambian material
genético. La recombinación genética resultante hace aumentar en gran
medida la variación genética entre la descendencia de progenitores que
se reproducen por vía sexual.
La recombinación genética está mediada por la aparición entre los dos
homólogos de una estructura proteica de 90 nm de diámetro llamada nódulo de recombinación. En él se encuentran las enzimas que medían en el proceso de recombinación.
Durante esta fase se produce una pequeña síntesis de ADN, que
probablemente está relacionada con fenómenos de reparación de ADN
ligados al proceso de recombinación.
- Diploteno
Los cromosomas continúan condensándose hasta que se pueden comenzar a
observar las dos cromátidas de cada cromosoma. Además en este momento
se pueden observar los lugares del cromosoma donde se ha producido la
recombinación. Estas estructuras en forma de X reciben el nombre quiasmas.
Cada quiasma se origina en un sitio de entrecruzamiento, lugar en el
que anteriormente se rompieron dos cromátidas homólogas que
intercambiaron material genético y se reunieron.
En este punto la meiosis puede sufrir una pausa, como ocurre en el caso de la formación de los óvulos humanos. Así, la línea germinal
de los óvulos humanos sufre esta pausa hacia el séptimo mes del
desarrollo embrionario y su proceso de meiosis no continuará hasta
alcanzar la madurez sexual. A este estado de latencia se le denomina dictioteno.
- Diacinesis
Esta etapa apenas se distingue del diplonema. Podemos observar los
cromosomas algo más condensados y los quiasmas. El final de la
diacinesis y por tanto de la profase I meiótica viene marcado por la
rotura de la membrana nuclear.
Durante toda la profase I continuó la síntesis de ARN en el núcleo. Al
final de la diacinesis cesa la síntesis de ARN y desaparece el nucléolo.
- Anotaciones de la Profase I
La membrana nuclear desaparece. Un cinetocoro se forma por cada cromosoma, no uno por cada cromátida, y los cromosomas adosados a fibras del huso
comienzan a moverse. Algunas veces las tétradas son visibles al
microscopio. Las cromátidas hermanas continúan estrechamente alineadas
en toda su longitud, pero los cromosomas homólogos ya no lo están y sus centrómeros y cinetocoros se encuentran separados.
Metafase I

El huso cromático aparece totalmente desarrollado, los cromosomas se
sitúan en el plano ecuatorial y unen sus centromeros a los filamentos
del huso.
Anafase I
Los quiasmas se separan de forma uniforme. Los microtúbulos del huso
se acortan en la región del cinetocoro, con lo que se consigue remolcar
los cromosomas homólogos a lados opuestos de la célula, junto con la
ayuda de proteínas motoras.
Ya que cada cromosoma homólogo tiene solo un cinetocoro, se forma un
juego haploide (n) en cada lado. En la repartición de cromosomas
homólogos, para cada par, el cromosoma materno se dirige a un polo y el
paterno al contrario. Por tanto el número de cromosomas maternos y
paternos que haya a cada polo varía al azar en cada meiosis. Por
ejemplo, para el caso de una especie 2n = 4 puede ocurrir que un polo
tenga dos cromosomas maternos y el otro los dos paternos; o bien que
cada polo tenga uno materno y otro paterno.

Telofase I
Cada célula hija ahora tiene la mitad del número de cromosomas pero
cada cromosoma consiste en un par de cromátidas. Los microtubulos que
componen la red del huso mitótico desaparece, y una membrana nuclear
nueva rodea cada sistema haploide. Los cromosomas se desenrollan
nuevamente dentro de la carioteca (membrana nuclear). Ocurre la citocinesis
(proceso paralelo en el que se separa la membrana celular en las
células animales o la formación de esta en las células vegetales,
finalizando con la creación de dos células hijas). Después suele ocurrir
la intercinesis,
parecido a una segunda interfase, pero no es una interfase verdadera,
ya que no ocurre ninguna réplica del ADN. No es un proceso universal, ya
que si no ocurre las células pasan directamente a la metafase II.

Meiosis II
La meiosis II es similar a la mitosis. Las cromátidas de cada
cromosoma ya no son idénticas en razón de la recombinación. La meiosis
II separa las cromátidas produciendo dos células hijas, cada una con 23
cromosomas (haploide), y cada cromosoma tiene solamente una cromátida.
Profase II

- Profase Temprana
Comienzan a desaparecer la envoltura nuclear y el nucleolo. Se hacen
evidentes largos cuerpos filamentosos de cromatina, y comienzan a
condensarse como cromosomas visibles.
- Profase Tardía II
Los cromosomas continúan acortándose y engrosándose. Se forma el huso
entre los centríolos, que se han desplazado a los polos de la célula.
Metafase II

Las fibras del huso se unen a los cinetocoros de los cromosomas.
Éstos últimos se alinean a lo largo del plano ecuatorial de la célula.
La primera y segunda metafase pueden distinguirse con facilidad, en la
metafase I las cromátides se disponen en haces de cuatro (tétrada) y en
la metafase II lo hacen en grupos de dos (como en la metafase mitótica).
Esto no es siempre tan evidente en las células vivas.
Anafase II

Las cromátidas se separan en sus centrómeros, y un juego de
cromosomas se desplaza hacia cada polo. Durante la Anafase II las
cromátidas, unidas a fibras del huso en sus cinetocoros, se separan y se
desplazan a polos opuestos, como lo hacen en la anafase mitótica. Como
en la mitosis, cada cromátida se denomina ahora cromosoma.
Telofase II

En la telofase II hay un miembro de cada par homólogo en cada polo.
Cada uno es un cromosoma no duplicado. Se reensamblan las envolturas
nucleares, desaparece el huso acromático, los cromosomas se alargan en
forma gradual para formar hilos de cromatina, y ocurre la citocinesis.
Los acontecimientos de la profase se invierten al formarse de nuevo los
nucleolos, y la división celular se completa cuando la citocinesis ha
producidos dos células hijas. Las dos divisiones sucesivas producen
cuatro núcleos haploide, cada uno con un cromosoma de cada tipo. Cada
célula resultante haploide tiene una combinación de genes distinta. Esta
variación genética tiene dos fuentes:
1.- Durante la meiosis, los cromosomas maternos y paternos se barajan, de modo que cada uno de cada par se distribuye al azar en los polos de la anafase I.
2.- Se intercambian segmentos de ADN.
1.- Durante la meiosis, los cromosomas maternos y paternos se barajan, de modo que cada uno de cada par se distribuye al azar en los polos de la anafase I.
2.- Se intercambian segmentos de ADN.
GAMETOGÉNESIS

La
gametogénesis es la formación de gametos por medio de la meiosis
a partir de células germinales. Mediante este proceso, el número de cromosomas
que existe en las células germinales se reduce de diploide
(doble) a haploide (único), es decir, a la mitad del
número de cromosomas que contiene una célula normal de la especie de que se
trate. En el caso de los hombres si el proceso tiene como fin producir espermatozoides
se le denomina espermatogénesis y se realiza en los testículos.
En caso contrario, si el resultado son ovocitos
se denomina ovogénesis y se lleva a cabo en los ovarios.
Este
proceso se realiza en dos divisiones cromosómicas y citoplasmáticas,
llamada primera y segunda división meiótica o simplemente meiosis I y meiosis
II. Ambas comprenden profase, prometafase, metafase, anafase, telofase y
citocinesis. Durante la meiosis I los miembros de cada par homólogo de
cromosomas se unen primero y luego se separan con el uso mitótico y se
distribuyen en diferentes polos de la célula. En la meiosis II, las cromátidas
hermanas que forman cada cromosoma se separan y se distribuyen en los núcleos
de las nuevas células. Entre estas dos fases sucesivas no existe la fase S
(duplicación del ADN).
La
meiosis no es un proceso perfecto, a veces los errores en la mitosis son
responsables de las principales anomalías cromosómicas. La meiosis consigue
mantener constante el número de cromosomas de las células de la especie para
mantener la información genética.
OVOGÉNESIS

La ovogénesis es el proceso de formación de los
gametos femeninos. Tiene lugar en los ovarios. Los ovogonios se ubican en los folículos ováricos,
crecen y tienen modificaciones; éstos llevan a la primera división meiótica que
da como resultado un ovocito primario (que contiene la mayor parte del
citoplasma) y un primer corpúsculo polar (su rol es llevarse la mitad de los
cromosomas totales de la especie).
Las dos células resultantes efectúan la meiosis
II, del ovocito secundario se forman una célula grande (que tiene la mayor
parte del citoplasma) y un segundo corpúsculo polar, estos se desintegran
rápidamente, mientras que la célula grande se desarrolla convirtiéndose en los
gametos femeninos llamados óvulos. El Gameto femenino queda estancado en
meiosis II, específicamente en Metafase II; si éste Gameto es fecundado, la
célula continúa Meiosis II para que sea Haploide. Al óvulo lo rodea una capa de
diferentes células, llamada folículo de Graaf.
La ovogénesis cuenta con diversas fases, las cuales
son:
- Proliferación: durante el desarrollo embrionario, las células germinales de los ovarios sufren mitosis para originar a los ovogonios.
- Crecimiento: en la pubertad crecen para originar los ovocitos de primer orden.
- Maduración: el ovocito del primer orden sufre meiosis.
La ovogénesis comienza antes del nacimiento y se
completa durante la vida reproductiva de la mujer, al ocurrir la fecundación.
ESPERMATOGÉNESIS

La espermatogénesis es el proceso de producción de los gametos masculinos
(espermatozoides) que tienen su producción en los testículos, específicamente
en los túbulos seminíferos. Dentro de este, destacan los siguientes procesos:
- Proliferación: las células germinales de los testículos sufren mitosis para que la cantidad de espermatogonios sea amplia.
- Crecimiento: las células germinales sufren su primera división meiótica para formar los llamados "espermatocitos 1". Luego sufren su segunda división meiótica, donde se forman los "espermatocitos 2".
- Maduración: los espermatocitos 2, que ya son haploides y de cromosomas simples, se les genera el flagelo y el acrosoma. A estos espermatocitos 2, luego de su transformación se les llama espermátida.
- Diferenciación: cada espermátida es diferente a otra por la variabilidad genética (crossing-over y permutación cromosómica).
En la espermatogénesis, por cada célula germinal se
producen cuatro espermátidas.
Gónadas
También llamadas órganos sexuales primarios funcionan
como glándulas mixtas en la medida que se producen hormonas y gametos. Los
órganos sexuales secundarios son aquellas estructuras que maduran en la
pubertad y que son esenciales en el cuidado y transporte de gametos, son rasgos
que se consideran de atracción sexual.
Los testículos son dos estructuras ovaladas que se
hallan suspendidas dentro del escroto mediante cordones espermáticos, son las
que producen semen y líquido testicular; su función endocrina es liberar
hormonas masculinas como la testosterona, quienes participaran en mantener los
caracteres sexuales masculinos.
Los ovarios son dos órganos con forma de almendra,
situados en los extremos de las trompas de Falopio, los ovarios son formados
aproximadamente cuando el feto hembra tiene 3 meses y cuando la mujer entra a
la pubertad los óvulos se van desarrollando. Su función endocrina es liberar
hormonas como la progesterona y estrógeno, las cuales
intervendrán en el ciclo ovárico.
Diferencias
entre espermatogénesis y ovogénesis
Espermatogénesis
- Se realiza en los testículos.
- Ocurre a partir de la espermatogonia.
- Cada espermatogonia da origen a cuatro espermatozoides.
- En la meiosis el material se divide equitativamente.
- Los espermatozoides se producen durante toda su vida.
- Se produce en el hombre.
- De un espermatocito I, se forman 4 espermios funcionales.
Ovogénesis
- Se realiza en los ovarios.
- Ocurre a partir de la ovogonia.
- Cada ovogonia da origen a un ovocito II el cual sólo en el caso de ser fecundado pasará a llamarse óvulo y a 2 cuerpos polares I y a un cuerpo polar II (sólo en caso de fecundación).
- En meiosis I no se divide el citoplasma por igual, quedando una célula hija (ovocito II) con casi todo el citoplasma.
- La mujer nace con un número determinado de folículos, aproximadamente 400.000.
- Se produce en la mujer.
- De un ovocito I, se forma un óvulo funcional.
Semejanzas
entre espermatogénesis y ovogénesis
- Ambos son subprocesos de la gametogénesis.
- Los dos producen gametos.
- En ambos se produce la meiosis.
- Los dos son procesos de la reproducción sexual en mamíferos.
- Ambos procesos se producen dentro de las gónadas.
- Los dos inician sus fases a partir de la meiosis.
Comparación
entre óvulos y espermatozoides
Ovocito II

- Más grande que el espermatozoide.
- Tiene vitelo (reserva nutritiva).
- No tiene movimiento.
- Sirve solo uno de cada célula germinal.
- Se produce en el ovario.
Espermatozoide

- Pequeño en comparación al ovocito II.
- No tiene reservas nutritivas.
- Se mueve por medio de su flagelo.
- Sirven cuatro de cada célula germinal.
- Se produce en el testículo.
No hay comentarios:
Publicar un comentario